August 12, 2016
Global crosstalk limits gene regulation
Scientists at the interface of biophysics, evolutionary biology and systems biology develop new framework to analyze effects of global crosstalk on gene regulation | interdisciplinary collaboration between three IST Austria research groups leads to publication in Nature Communications
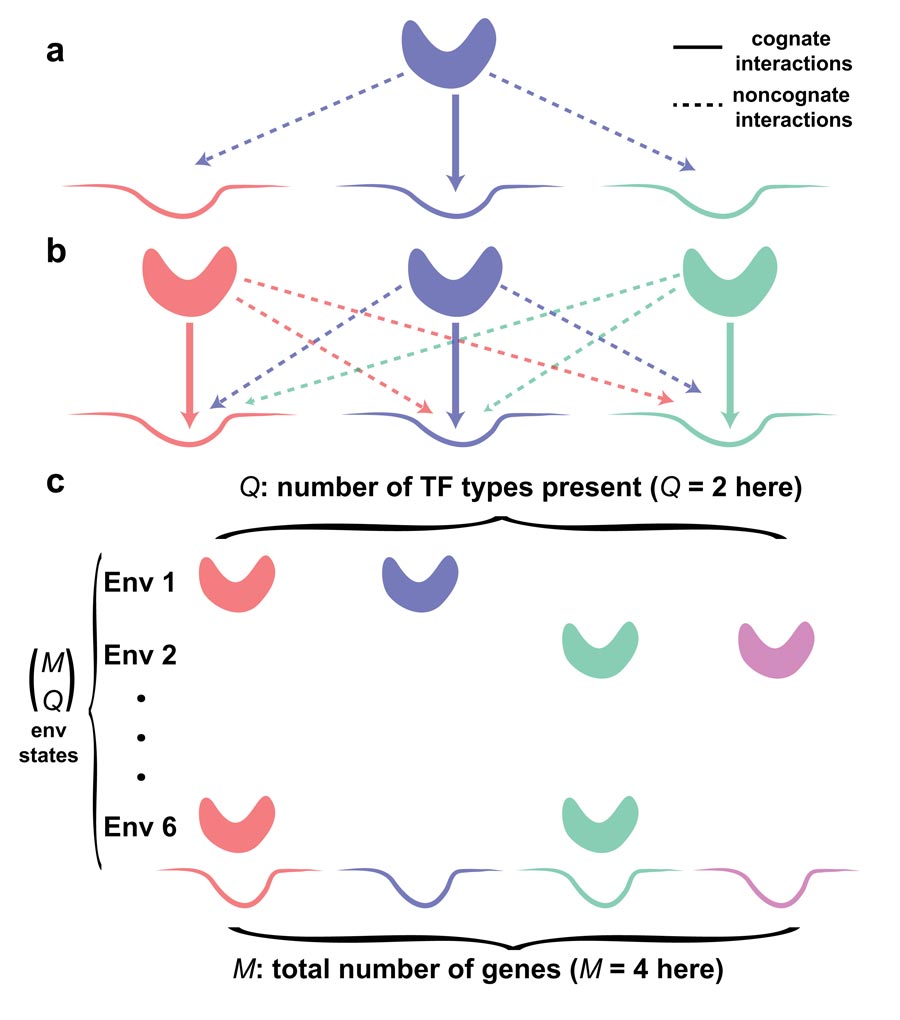
Molecular recognition is fundamental to transcriptional regulation, the primary mechanism by which cells control gene expression. The specificity of this regulation originates in the binding interactions between special regulatory proteins, called transcription factors (TFs), and short regulatory sequences on the DNA, called binding sites. Although each type of TF preferentially binds certain regulatory DNA sequences, evidence shows that this binding specificity is limited, and that TFs bind other noncognate targets, too. If these sites happen to be regulatory elements of other genes, noncognate binding not only depletes TF molecules, but could also actively interfere with gene regulation. This suggests that the crosstalk problem is global.
In “Intrinsic limits to gene regulation by global crosstalk”, a research paper published on August 4 in Nature Communications, ISTFellow Tamar Friedlander, PhD student Roshan Prizak, and Professors Calin Guet, Nick Barton and Gasper Tkacik construct a biophysical model for crosstalk in transcriptional regulation that accounts for all cross-interactions between regulators and their binding sites. They identify the parameters that have a major influence on crosstalk severity. While some of these parameters are difficult to estimate, they show that there exists a lower bound to crosstalk with respect to these parameters. This implies the existence of a “crosstalk floor,” which cannot be overcome even if TF concentrations were optimally adjusted by the cell and compensated for sequestration at noncognate sites.
Although most of the biophysical constraints have been understood at the level of individual genetic regulatory elements, the researchers find that crosstalk is special: while it originates locally due to biophysical limits to molecular recognition, its cumulative effect only emerges globally. At the level of a single genetic regulatory element, crosstalk can always be avoided by increasing the concentration of cognate TFs or introducing multiple binding sites in the promoter. It is only when these same cognate TFs act as noncognate TFs for other genes, or that new binding sites in the promoter drastically increase the number of noncognate binding configurations, that crosstalk constraints become clear.
